转自:地质地球所 发布日期:2022-5-14
在真核生物中硅藻、硅质海绵、放射虫等的生物矿化及其遗骸埋藏构成了显生宙以来硅循环的重要过程,它们矿化被认为是海水可溶性硅浓度快速降低的主控因素之一。长期以来,硅的生物矿化被认为是许多真核生物的专属特性。在真核生物出现前的早期地球上硅质岩被认为主要是化学沉淀引起的。近年来,一些研究显示:原核生物可能具有硅化能力。如果原核生物能够参与硅的生物矿化,地球早期的硅循环过程将需要被重新评估。
近日,中国科学院地质与地球物理研究所李金华研究员与潘永信院士生物地磁学团队联合西北大学、巴黎第六大学、加拿大国家同步辐射中心、中科院生物物理所、物理所和厦门大学等国内外多个单位科研人员,利用先进电子显微学和同步辐射技术观测,发现原核生物趋磁细菌中一类硝化螺旋菌门趋磁球菌,在细胞内合成亚微米尺寸的非晶质二氧化硅(SiO2)球粒沉淀,平均直径约为0.53 ± 0.34 μm,约占细胞总质量的~10.9%,与硅藻细胞中二氧化硅含量相当。该项研究为进一步探索地球演化早期的硅循环的成因提供了新思路。

这幅图展示了该研究新发现的趋磁球菌WYHC-5及其亚细胞结构。
(A)高角环形暗场扫描透射电子显微镜图像展示WYHC-5在细胞内合成纳米尺寸的磁铁矿链和亚微米尺度的二氧化硅球状颗粒;
(B)聚焦离子束-扫描电镜三维重构图展示WYHC-5 胞内的二氧化硅、液泡和磁小体链的空间构象;
(C)高压冷冻替代细胞的透射电镜超薄切片图像展示WYHC-5细胞内的亚微米尺寸液泡和纳米尺寸类羧酶体结构;
(D)高压冷冻替代细胞的聚焦离子束-扫描电镜三维重构图像展示WYHC-5 胞内的液泡、类羧酶体和磁小体的空间构象
硅元素与生物硅化
硅在宇宙中的储量排在第八位。在地壳中,它是第二丰富的元素,构成地壳总质量的26.4%,仅次于氧(49.4%)。在现代工业和高科技产业中,硅的应用极其广泛,主要用来制作高纯半导体、耐高温材料、光导纤维通信材料、有机硅化合物和合金等。在自然界,硅通常以二氧化硅(SiO2)或硅酸盐(Silicates)的形式,广泛存在于土壤和岩石中。大陆硅酸盐的风化作用可以移除大气CO2,将硅和溶解的无机碳迁移至海洋,被认为是调节二氧化碳水平,控制地球气候的关键机制。
硅是许多高等生物必须的微量或常量元素。比如,在人体中,硅参与软骨和结缔组织的形成。在高等植物中,硅以非晶质二氧化硅(SiO2·H2O)的形式积累,从而提高植物茎秆的硬度以及对干旱、盐胁迫、紫外辐射和病虫害等的抗性。硅藻、放射虫和硅质海绵甚至以硅作为营养元素,从环境中摄取大量可溶性硅,并将其转化成二氧化硅,用它构建细胞骨架或者躯壳。这些生物的生命活动及其硅质遗骸的埋藏构成了显生宙以来全球硅-碳循环的主要环节,被认为是海水可溶性硅浓度快速降低的主控因素。以硅藻为例,它们是一类真核单细胞藻类,是海洋中最常见的浮游生物,通过光合作用将大气中CO2转化为颗粒有机碳,通过生物硅化作用将水中的可溶性硅转化为细胞骨架二氧化硅,经沉降运移至深海储藏,贡献了约40%的全球海洋初级生产力,对大气CO2含量有重要调节作用,同时可能造成海水中可溶性硅浓度从上千μM降至现今的~10 μM。
在早期的认识中,原核生物被认为不能通过生物控制矿化作用沉积二氧化硅。2001年美国炭疽攻击事件中,美国联邦调查局(FBI)发现通过邮件递送到美国的炭疽芽孢粉末被二氧化硅包裹。这究竟是生物矿化导致的,还是将制备后的芽孢用二氧化硅包裹以提高其稳定性、增强其分散性?这个问题曾引发了近十年的争论。2010年,日本广岛大学研究人员证实:枯草芽孢杆菌在其芽孢(内生孢子,一种细菌的休眠体)外壁沉淀形成一层二氧化硅薄膜,可以提高芽孢抗酸腐蚀和紫外辐射的能力。2012年,美国石溪大学研究人员发现聚球藻属的蓝细菌能从周围水体中摄取和富集大量的硅。这些研究引发了一个问题:新元古代硅质海绵出现之前,生物,尤其是微生物是否参与了地球上硅的生物地球化学循环?
趋磁细菌与生物矿化
趋磁细菌起源于太古代并广泛分布于各种水体环境中,它们合成磁小体,感知地磁场,在地质记录中形成磁小体化石,因而是生物矿化、生物地磁学和古地磁学研究的理想模式系统。通过跨学科交叉研究,中国科学院地质与地球物理研究所生物地磁学研究团队在近期的研究发现,除了磁铁矿或胶黄铁矿的晶体颗粒,不同趋磁细菌类群可以在细胞内合成不同的有机颗粒物和生物矿物:如α-变形菌纲趋磁细菌可以在细胞内合成聚β-羟基丁酸(PHB),脱硫菌门趋磁细菌在细胞内合成多聚偏磷酸,硝化螺菌门和暂定杂食菌门的趋磁细菌在细胞内合成单质硫颗粒。此外,有些趋磁细菌既能合成多聚偏磷酸又能合成单质硫颗粒。一些特殊种类的趋磁细菌可以在细胞内同时合成其它类型的矿物,如磷酸铁、磷酸钙、碳酸钙,甚至一些富含Ca/Ba的未知矿物。自然环境中的趋磁细菌死亡裂解后,这些由趋磁细菌合成的复杂多样的有机颗粒物和生物矿物也可以像磁小体一样,有可能被埋藏并保存在沉积物中。因此,除了为沉积物和沉积岩提供磁性矿物源,趋磁细菌还为沉积环境贡献有机质(细胞碎屑和PHB)、单质硫、二氧化硅、磷酸盐和碳酸盐等生物矿物。而且,趋磁细菌的代谢产物,如硫酸盐还原过程中产生的硫化氢,多聚偏磷酸降解后产生的磷酸根,可以通过复杂的化学反应以单质硫的形式在沉积物种沉淀,或者通过微生物催化的氧化还原过程以黄铁矿或者磷酸铁的形式沉淀。
趋磁细菌中首次发现二氧化硅球粒沉淀
2017年,中国科学院地质与地球物理研究所生物地磁学研究团队在研究趋磁细菌生物矿化和磁学性质的过程中,从陕西西安未央湖沉积物中发现了一类独特的硝化螺旋菌门趋磁球菌WYHC-5(视频1)在细胞内合成亚微米尺寸的非晶质二氧化硅球粒沉淀。这是首次发现了原核生物胞内生物控制矿化二氧化硅的新机制。
详细的实验观测如下:
(1)显微学分析揭示WYHC-5的细胞平均直径为3.58 ± 0.42 μm,基于16S rRNA基因序列的系统发育分析表明,WYHC-5属于古老的硝化螺旋菌门趋磁细菌。这类趋磁球菌能在细胞内合成近百个弯曲的子弹头形磁铁矿磁小体,其平均长度为70.4 ± 27.0 nm,平均宽度为27.5 ± 6.3 nm。磁小体在细胞内被组装成数条松散的链束状结构。除磁小体外,WYHC-5还能在细胞合成大量球状颗粒(图1)。
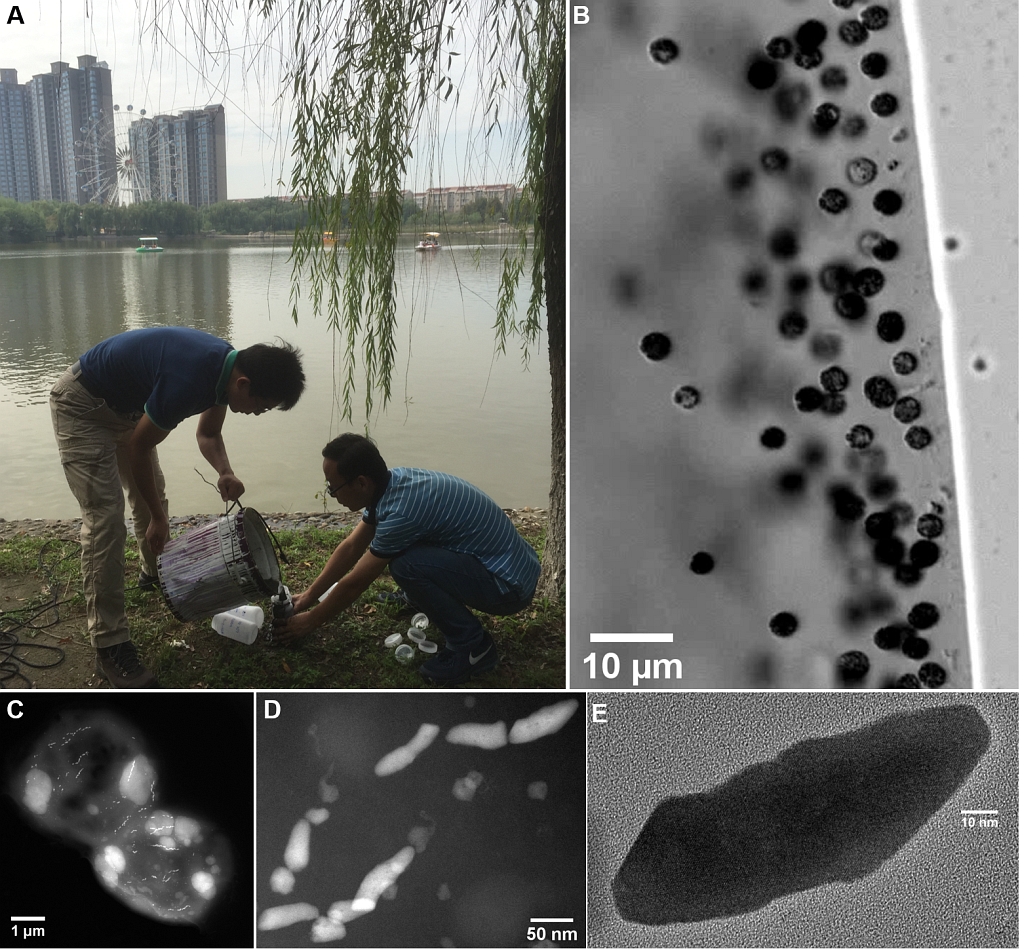
图1 陕西西安未央湖沉积物采样和趋磁球菌WYHC-5的实验室发现。
(A)湖底沉积物样品采集(李金华研究员和硕士研究生张衡);
(B)光学显微镜观察发现趋磁球菌WYHC-5;
(C)WYHC-5的透射电镜照片显示细胞内纳米尺寸的磁小体链和亚微米尺寸的球状颗粒物;
(D)WYHC-5细胞内的子弹头形磁小体;
(E)单个磁铁矿(Fe3O4)磁小体颗粒的高分辨透射电镜图像
(2)研究团队综合采用先进的透射电镜(HAADF-STEM EDXS,TEM-EESL)和同步辐射扫描透射X-射线显微镜(STXM)技术,对WYHC-5细胞内颗粒物的结构和成分进行了系统研究(图2)。结果发现:(i)WYHC-5磁小体富含Fe和O元素,为磁铁矿相,约占细胞总质量的~0.51%;(ii)球状颗粒物分两类,尺寸较大的富含Si和O元素,为非晶质的二氧化硅颗粒,平均直径约为0.53 ± 0.34 μm,约占细胞总质量的~10.9%;(iii)尺寸较小的颗粒物富含S元素,为具有线状结构的单质硫(S0)颗粒,平均直径约为0.38 ± 0.23 μm,约占细胞总质量的~3.2%。
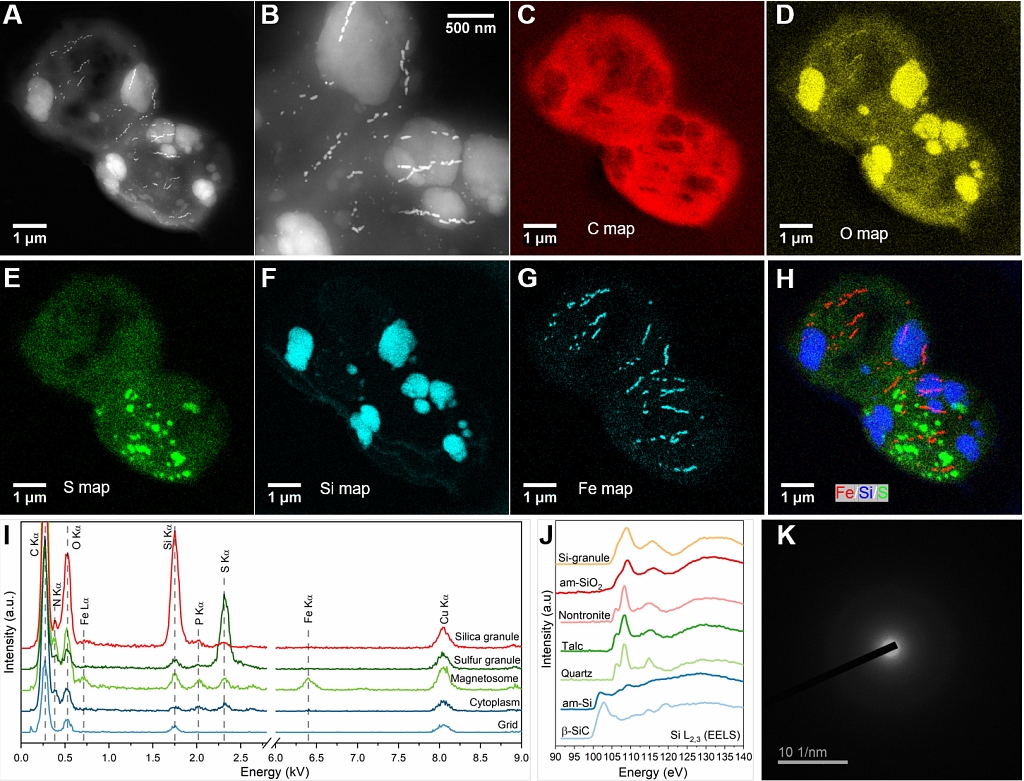
图2 透射电镜技术分析WYHC-5细胞内颗粒物成分和结构。
(A)两个代表性WYHC-5细胞的高角环形暗场扫描透射电镜图像(HAADF-STEM);
(B)WYHC-5细胞的局部放大图(HAADF-STEM);
(C-H)不同元素分布的伪彩图(STEM-EDXS elemental maps);
(I)WYHC-5细胞内不同感兴趣区域的电子能量色散谱图(EDXS);
(J)WYHC-5细胞内的二氧化硅颗粒的透射电镜电子能量损失谱图(TEM-EESL)及其与其它硅化合物的比较;
(K)WYHC-5细胞内二氧化硅的透射电镜选区电子衍射花样(SAED)
(3) 为了进一步确定细胞内不同颗粒物的空间构象,研究团队综合采用高压冷冻替代超薄切片透射电镜(High-pressure freeze and freeze substitution for ultramicrotomy and TEM)和聚焦离子束-扫描电镜三维重构(FIB-SEM tomography)技术,对WYHC-5细胞进行了更为精细的逐层切片研究(图3,视频2和视频3)。研究发现,除了磁小体、二氧化硅和单质硫颗粒物外,WYHC-5细胞内还含有大量亚微米尺寸的具有刚性膜包裹的液泡结构(平均尺寸为216.6 ± 64.7 nm),以及大量平均尺寸为107.6 ± 14.8 nm大小的类羧酶体结构。

图3 聚焦离子束-扫描电镜三维重构技术分析WYHC-5亚细胞结构。
(A)一个干燥的WYHC-5细胞的聚焦离子束-扫描电镜切面图像,展示细胞内的液泡、二氧化硅和单质硫颗粒,以及磁小体的空间构象;
(B)一个高压冷冻替代的WYHC-5细胞的透射电镜超薄切片图像,展示细胞内的液泡和类羧酶体结构;
(C)局部放大的透射电镜图像,展示磁小体膜和类羧酶体的精细结构;
(D)一个完整的WYHC-5细胞(高压冷冻替代处理后)的聚焦离子束-扫描电镜三维重构图像。高压冷冻替代预处理有利于呈现细胞内液泡、磁小体膜和类羧酶体的精细结构,但会造成二氧化硅和单质硫颗粒的溶解
(4)为了进一步确认二氧化硅生物矿化在趋磁细菌类群是否存在普遍性,研究团队进一步筛查了17种趋磁细菌,依据HAADF-STEM-EDXS化学成分分析, 从中发现了另外三株趋磁球菌WYHC-1(发现自西安未央湖)、YQC-1(发现自天津于桥水库)和MYC-4(发现自北京密云水库)均能在细胞内合成二氧化硅颗粒,这三种趋磁细菌从系统发育分类上属于远离硝化螺菌门的暂定分类单元的η系变形菌纲。这说明,不是所有的趋磁细菌都有生物沉硅能力,但二氧化硅的生物矿化至少在趋磁细菌系统内仍具有一定的分布。
(5)原核生物中硅生物合成二氧化硅(图4)。本研究首次揭示出原核生物中趋磁细菌的生物沉硅现象。枯草芽孢杆菌将二氧化硅沉积在芽孢外壁,而聚球藻属蓝细菌形成的是含镁硅酸盐,其生物沉硅发生在胞外还是胞内仍有待确认。与前两者不同,趋磁细菌通过主动跨膜运输,将周围环境中低浓度的可溶性硅摄入细胞内,浓度提升近3500倍(依据水体中可溶性硅浓度测量值与WYHC-5细胞内二氧化硅的平均质量换算),并矿化形成二氧化硅球状颗粒物,因此代表原核生物中一种全新的、高效的二氧化硅生物沉淀模式。
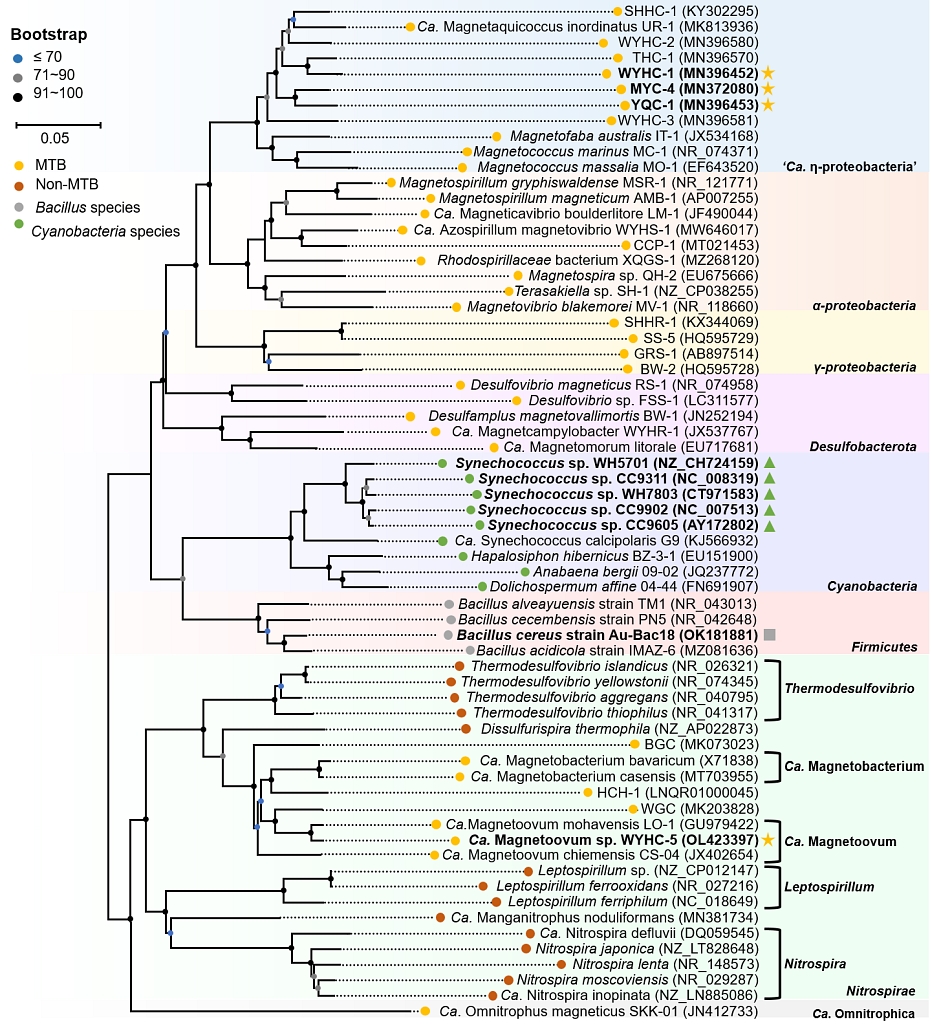
图4 具有生物沉硅能力的原核生物的系统发育树。黄色五角星代表能在细胞内矿化合成二氧化硅颗粒的趋磁细菌,绿色三角和灰色方框分别代表具有生物沉硅能力的聚球藻属蓝细菌和枯草芽孢杆菌
趋磁细菌是否有硅转运蛋白系统和沉硅酶?
趋磁细菌是一类能在细胞内合成纳米磁性晶体颗粒(也被称为磁小体,化学成分为Fe3O4或Fe3S4)的原核生物。磁小体多呈链状排列,作为细胞的小磁针,可以感知地磁场,使其沿磁力线方向游泳,在水体中“有氧-缺氧过渡带”上下穿梭,从而迅速找到最适合生存的微环境。问题是,除了磁小体,趋磁细菌为什么还要在细胞内沉积大量的二氧化硅?其分子机制是什么?这种在一个细胞内合成多种矿物的生理学和生态学意义是什么?为了弄清上述问题,研究团队对WYHC-5细胞进行了基因组测序分析,并与其它硅代谢相关生物的基因组开展的生物信息学比较分析。结果如下:
(1)与研究团队近期在密云水库趋磁大杆菌MYR-1中的研究一致,WYHC-5基因组中具有成套的磁小体矿化合成相关基因,完整的硝酸盐还原和亚硝酸盐氧化酶基因,完整的硫氧化(S2-→S0→SO32-→SO42-)和硫酸盐还原(SO42-→SO32-→S2-)关键酶基因,碳固定的自养生长和有氧呼吸的异养生长相关的关键酶基因,以及编码羧酶体的关键基因。据此推断,WYHC-5采用与趋磁大杆菌MYR-1相似的策略,利用地磁场的定向作用,在水生环境的缺氧环境和有氧环境中上下穿梭:在OATZ下部的无氧环境中,它利用硝酸盐将硫化氢氧化成单质硫以颗粒的形式沉积在细胞内(S2-→S0)后,向上游泳到OATZ上部的有氧环境中将存储的硫被彻底氧化成硫酸盐(S0→SO32-→SO42-),并自养生长,同时吸收硝酸根并将其存储在液泡中,然后,“怀揣”硝酸盐再次向下游泳到OATZ下部的无氧环境,利用硫酸盐进行异养生长或利用硝酸盐作为氧化剂,将硫化氢氧化成单质硫。这样,WYHC-5在细胞内沉淀非晶质二氧化硅颗粒的生理学意义可能在于:通过增加细胞的重量而调整细胞的浮力,从而有利于WYHC-5在水体中上下穿梭时保持稳定性。另一方面,WYHC-5利用HCO3-进行自养生长时,会消耗细胞内大量的H+,从而造成局部环境中pH升高,而二氧化硅矿化过程可以释放H+,从而为细胞的自养生长提供pH缓冲剂的作用。
(2)研究团队暂时没有从WYHC-5的基因组中发现与其它生物的硅转运蛋白序列相似的硅转运蛋白,这暗示WYHC-5细胞内可能存在独特的、但有待进一步鉴定的硅转运系统。然而,研究团队在WYHC-5基因组中发现了一个潜在的沉硅酶基因 (silicatein-1),它编码一个248氨基酸长度的蛋白质,与硅质海绵和鼠妇中已经鉴定的沉硅酶(Silicatein)具有约38.3%的蛋白质序列同源性。蛋白质结构解析和分子建模表明,WYHC-5的沉硅酶拥有一个由“半胱氨酸-组氨酸-天门冬氨酸”组成的催化结构域和一个丝氨酸簇,这是硅质海绵沉硅酶催化二氧化硅形成和结构组装的核心催化域。因此,我们推测silicatein-1沉硅酶可能负责WYHC-5细胞中的球状二氧化硅颗粒的形成(图5)。
(3)枯草芽孢杆菌、聚球藻属蓝细菌和趋磁细菌的硅代谢相关基因也各不相同。生物信息学分析表明,趋磁球菌WYHC-5的沉硅酶与硅质海绵沉硅酶序列高度同源,而枯草芽孢杆菌的芽孢壁蛋白(CotB1)似乎与硅藻的沉硅蛋白Silacidian A同源。而且,不同种类的枯草芽孢杆菌和聚球藻属蓝细菌似乎含有各自的硅转运系统。生物沉硅能力在三大类细菌类群中的“随机”分布也进一步说明,原核生物的生物沉硅可能具有古老起源,但在后期演化过程,大多数微生物选择丢掉了这一功能。另一种可能是,原核生物的生物沉硅在不同的微生物类群中具有多起源,或者这种生物沉硅能力干脆就是近代才发生的,通过基因的水平转移从不同类型的高等生物中向原核生物中进行传递。未来,确定原核生物的生物沉硅能力形成的时间,将为确定其是否参与早期地球硅循环提供关键证据。
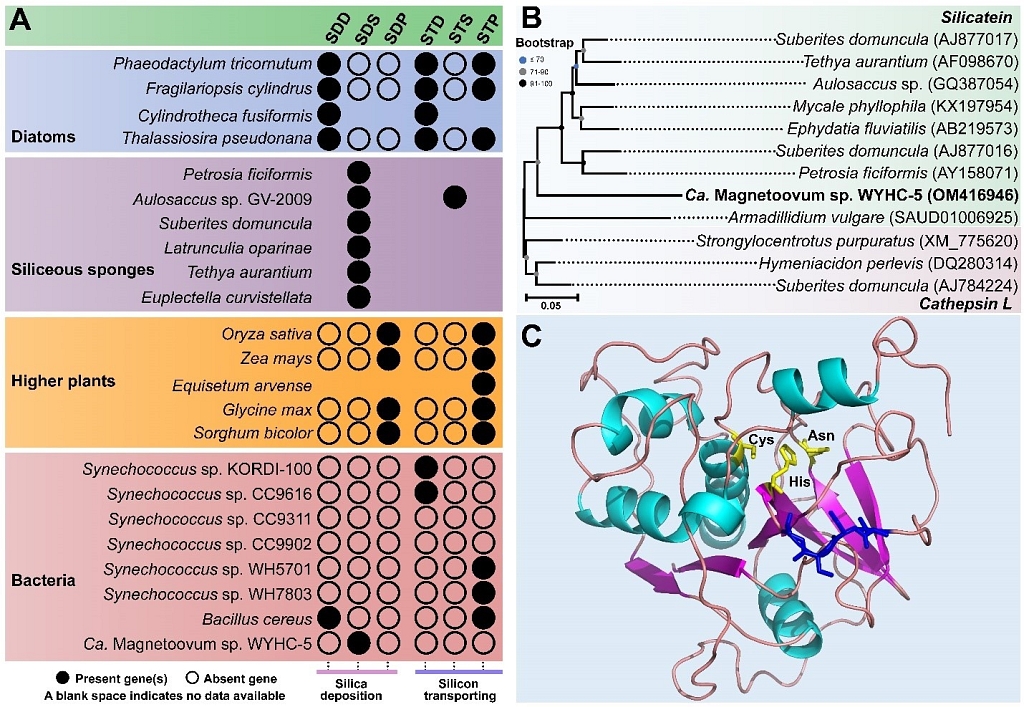
图5 趋磁球菌WYHC-5细胞中硅代谢相关基因及其功能预测。
(A)硅藻、硅质海绵、高等植物和细菌中的硅沉积(SD)和硅转运(ST)基因及其分布;
(B)沉硅酶(Silicatein)和组织蛋白酶L(Cathepsin L)基因的聚类分析;
(C)WYHC-5细胞沉硅酶的蛋白质分子建模和结构分析
“趋磁细菌驱动有氧-缺氧过渡界面物质和能量循环”工作模型
根据一系列的研究,研究团队提出了一个“趋磁细菌驱动有氧-缺氧过渡界面(OATZ)物质和能量循环”的工作模型:在地磁场的导向作用下,趋磁细菌在水体OATZ环境中上下高效穿梭,并通过生物矿化作用和生理代谢活动,将OATZ底部的厌氧环境与其上部的有氧或微氧环境联动起来,从而驱动C、N、P、S、Fe、Si等元素的生物地球化学循环,并参与一些矿物的生物形成或转化(图6)。因此,研究趋磁细菌为认识地球早期“地磁场-环境-生命”协同演化提供了一个新的途径。
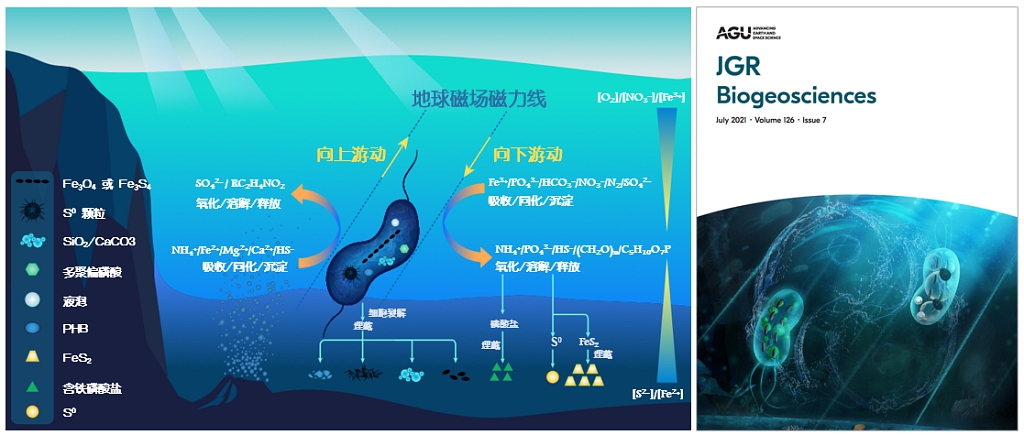
图6 (左)趋磁细菌利用地磁场的导向作用驱动地球水生环境中有氧-缺氧过渡带元素生物地球化学循环工作模型;(右)结合中国太极文化理念,为美国地球物理学会期刊JGR封面艺术化的工作模型图(Li et al., JGR, 2020; Li et al., JGR, 2021)
相关成果近期发表于Science Advances上(李金华*, 刘沛余, Menguy Nicolas, 张兴亮, 王建, Benzerara Karim, 冯连君, 孙磊, 郑越, 孟繁琦, 谷林, Leroy Eric, 郝佳龙, 储雪蕾, 潘永信. Intracellular silicification by early-branching magnetotactic bacteria[J]. Science Advances 2022, 8(19), eabn6045. DOI: 10.1126/sciadv.abn6045)(原文链接)。该项目研究受中国国家自然科学基金重点国际(地区)合作研究项(41920104009)、国家自然科学基金重大项目课题(41890843)和国家自然科学基金创新研究群体项目(41621004)资助。
| Copyright © 古地磁与年代学实验室 京ICP备05029136号 地址: 北京市北土城西路19号古地磁实验室 邮编:100029 电话:010-82998366 |

